





Abstract
Next generation biofuels including longer-chain alcohols such as butanol are attractive as renewable, high-energy fuels. A barrier to microbial production of butanols is the increased toxicity compared to ethanol; however, the cellular targets and microbial defense mechanisms remain poorly understood, especially under anaerobic conditions used frequently in industry. Here we took a comparative approach to understand the response of Saccharomyces cerevisiae to 1-butanol, isobutanol, or ethanol, across three genetic backgrounds of varying tolerance in aerobic and anaerobic conditions. We find that strains have different growth properties and alcohol tolerances with and without oxygen availability, as well as unique and common responses to each of the three alcohols. Our results provide evidence for strain-by-alcohol-by-oxygen interactions that moderate how cells respond to alcohol stress.
The increasing interest in sustainable energy has propelled the use of genetically engineered microbes to produce fuels from non-food plant biomass. While past research has focused on ethanol production from renewable feedstock, longer-chain alcohols such as butanol and isobutanol are more attractive as next generation biofuels due to their higher energy content and compatibility with existing infrastructure for gasoline distribution. In addition, butanol isoforms are less hygroscopic and have a low freezing point, which allows for blending up to 85% with gasoline (Agency, 2011) (Dürre, 2007, Fortman et al. 2008). These molecules can also be modified through chemical processes to generate even more powerful sources of energy such as jet fuels (Taylor et al. 2010).
Although the industrial microbe Saccharomyces cerevisiae does not produce significant amounts of four-chain alcohols natively, engineering S. cerevisiae for higher-titer butanol production is underway (Avalos et al. 2013, Chen & Liao, 2016, Liu et al. 2017). Production of 1-butanol in yeast is enabled by introduction of genes involved in the acetone-butanol-ethanol (ABE) clostridial fermentation, which enables the conversion of acetyl-CoA to 1-butanol (Steen et al. 2008, Lian et al. 2014, Generoso et al. 2015, Swidah et al. 2015, Schadeweg & Boles 2016). Deletion of the alcohol dehydrogenase gene ADH1 can also enhance native 1-butanol production (Si et al. 2014). In contrast, production of isobutanol has been achieved by rerouting valine metabolism to produce isobutanol in mitochondria or the cytosol (Chen et al. 2011, Brat et al. 2012, Kondo et al. 2012, Matsuda et al. 2013, Hammer & Avalos 2017), with highest efficiency when the whole pathway is engineered in the same cellular compartment (Avalos et al. 2013, Park et al. 2016). Although there have been successes in engineering S. cerevisiae to produce these molecules, yields are still low, making the production of next generation biofuels economically limiting at this time.
Engineering improved end-product tolerance in the host is another important consideration, since alcohol toxicity likely limits production of these fuels (Fischer et al. 2008, Dunlop, 2011, Generoso et al. 2015). Many studies have focused on the mechanism of inhibition caused by ethanol (Meaden et al. 1999, Alexandre et al. 2001, Aguilera et al. 2006, Fujita et al. 2006, Hu et al. 2007) and identified engineering strategies that increased ethanol tolerance (Hu et al. 2007, Lewis et al. 2010, Swinnen et al. 2012, Hubmann et al. 2013, Lam et al. 2014, Zyrina et al. 2017). However the inhibitory mechanisms of longer-chain alcohols in S. cerevisiae are less well understood. Longer chain alcohols are known to cause significantly more membrane damage than ethanol (Gray & Sova, 1956, Liu & Qureshi, 2009, Huffer et al. 2011), disrupting pH balance and inhibiting important membrane proteins such as ATPases and glucose transporters (Paterson et al. 1972, Grisham & Barnett, 1973, Ingram, 1976, Bowles & Ellefson, 1985, González-Ramos et al. 2013). In addition, these alcohols are known to induce the accumulation of misfolded proteins (Dunlop, 2011, Ghiaci et al. 2013, González-Ramos et al. 2013, Navarro-Tapia et al. 2016). It is largely unclear how alcohol tolerance is affected under anaerobic conditions, which is an important consideration given that many industrial processes are performed anaerobically. Both anaerobiosis and alcohols affect membranes, but in different ways. The absence of oxygen prevents the production of sterols and unsaturated fatty acids and thus alters membrane composition and fluidity (Wilcox et al. 2002, Rosenfeld & Beauvoit 2003), while alcohols target membrane integrity directly (Ingram, 1976, Ingram, 1986, Mishra & Prasad, 1989, Alexandre et al. 1994). In turn, membrane composition influences alcohol tolerance (Mannazzu et al. 2008, Henderson et al. 2011, Henderson et al. 2013, Archana et al. 2015), although the optimal membrane composition and the mechanisms of tolerance remain unclear (Huffer et al. 2011, Henderson & Block 2014). Other studies have linked respiration, protein folding, and protein degradation to aerobic butanol tolerance (Ghiaci et al. 2013, González-Ramos et al. 2013, Crook et al. 2016). Exploring how hypoxia modifies the inhibitory effects of alcohols has important industrial applications. But an additional important consideration is that different strains may vary in their responses and the mechanisms they use to survive anaerobic stresses.
Here, we used genetic and genomic approaches to explore the differences in alcohol response in the presence and absence of oxygen, across multiple genetic backgrounds of yeast. Our past strategy compared stress responses across natural strains with varying tolerance to identify primary targets of industrial stressors and defense mechanisms employed by tolerant strains (Lewis et al. 2010, Sardi et al. 2016, Sardi et al. 2018). Here we compared the transcriptome responses to 1-butanol, isobutanol, and ethanol in both aerobic and anaerobic conditions, across three strain backgrounds with varying tolerances, using RNA sequencing (RNA-seq). The strains showed significant differences in their response to anaerobic growth alone and in their tolerance and response to alcohols under aerobic and anaerobic conditions. Comparing and contrasting across alcohols and aerobic conditions revealed unique responses to ethanol vs. longer-chain alcohols and synergistic effects between alcohols and oxygen depletion. This study therefore revealed important genotype by environment by environment interactions that affect the stress response and alcohol tolerance under different conditions. Together, these results expand our knowledge of alcohol responses for future engineering strategies while presenting important information about how environments and genotype interact in a stress response.
Methods
Strains and growth conditions
Strains used in this study and their phenotypes are listed in Table S1. Three strains were selected for more detailed analysis: West African strain NCYC3290 (WA), mosaic (i.e., admixed from pure lineages) isolate from Illinois, USA IL01 (courtesy Justin Fay) and a mosaic strain isolated from papaya, Y7568 (PAP). The concentrations of ethanol, butanol, and isobutanol were selected as the maximal tested doses that allowed cell growth in all three strains. Unless otherwise noted, cells were grown in shaking flasks at 30°. Rich lab medium (YPD: 1% yeast extract, 2% peptone, 2% dextrose) was used as the base medium for all growth conditions. Anaerobic growth was performed in an anaerobic chamber maintained at O2 < 25 PPM, using culture flasks with a stir bar to maintain cell suspension. Phenotypes for Figure 1B-C were measured using 1% 1-butanol, 1.2% isobutanol, and 5% ethanol.
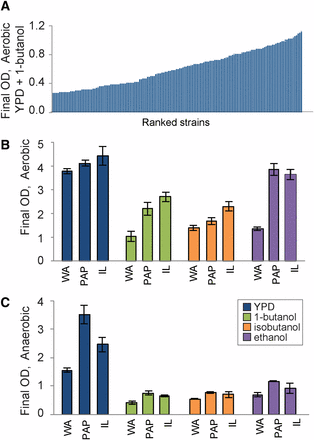
Strain-specific tolerance of alcohols varies with oxygen. (A) Representative final OD600 measurements of 165 strains grown in YPD with 2% butanol. Strains are sorted by final OD (Table S1). (B-C) West African strain NCY3290 (WA), mosaic strains Y7568 (PAP) and IL01 (IL) were grown in YPD with and without added alcohols. Final OD600 of strains grown (B) aerobically 10 h or (C) anaerobically for 11 h in YPD, YPD + 1% butanol, 1.2% isobutanol, or 5% ethanol. Data represent the average and standard deviation of 3 replicates.
Phenotyping
High-throughput phenotyping of the 165-strain Saccharomyces cerevisiae collection was performed in 96-well plates (NUNC, Thermo Scientific, Rockford, IL). Briefly, 10 µl of thawed frozen stock of cells was used to inoculate a 96-well plate containing 190 µl of YPD media. Plates were sealed with breathable tape (AeraSeal, Sigma, St. Louis, MO), covered with a lid and incubated at 30° while shaking for 24 hr, at which time a new subculture was generated by inoculating 190 µl of YPD with 10 µl of the previous culture and grown to log phase for 6 hr. 10 µl of this log-phase culture was inoculated into 190 µl of YPD + 2% butanol, plates were then sealed with an aluminum foil seal (Cryostuff, Vienna, VA, USA) to minimize butanol evaporation, grown with shaking for 24 hr, followed by measurement of the final OD600 using the Tecan M200 Pro microplate reader (Tecan Systems, Inc., San Jose, CA). The average of four biological replicates was calculated to represent tolerance to butanol. Single-strain phenotyping was performed in shaker flasks at 30C under defined media conditions.
Oxygen utilization rate measurement
Strains were grown in rich lab media (YPD) for three generations and kept in log phase before oxygen measurements. A microoptode oxygen probe (Unisense, Denmark) was used to measure dissolved oxygen over time, measuring every 15 sec for 3 min. Each measurement was normalized to underlying OD600 and oxygen consumption was taken as the slope of measured oxygen over time.
Transcriptome profiling
Strains were grown at 30° with shaking to mid-log phase for seven generations in YPD. The culture was then used to inoculate YPD, YPD + 0.8% 1-butanol, YPD + 1% isobutanol, YPD + 4% ethanol (or YPD + 3% ethanol for anaerobic conditions, since WA did not grow well anaerobically on 4% ethanol), grown for three generations and collected during log phase by centrifugation. Collections were performed in duplicate on different days, such that replicate pairs could be analyzed using paired statistics. Analyzing all 12 samples together with a multi-factorial linear model provided additional statistical power (see below). RNA was processed as previously described (Sardi et al. 2016). Briefly, RNA was extracted by hot phenol lysis (Gasch 2002). Total RNA was DNase-treated at 37° for 30 min with TURBO DNase (Life Technologies, Carlesbad, CA), followed by RNA precipitation at -20° in 2.5M LiCl for 30 min. rRNA depletion and library generation was constructed by the University of Wisconsin-Madison Biotechnology Center, via the TrueSeq Stranded Total RNA Sample Preparation Guide (Rev.C) using the Illumina TrueSeq Stranded Total RNA (yeast) kit (Illumina Inc., San Diego, California, USA). Libraries were standardized to 2 µM. Cluster generation was performed using standard cluster kilts (vs) and the Illumina cluster station. Single-end 100 bps reads were generated on an Illumina HiSeq2500 sequencer. All sequencing data are available in the NIH GEO database under accession number GSE118069.
Reads were processed with Trimmomatic (Bolger et al. 2014) and mapped to reference genome S288c (NC_001133, version 64 (Engel et al. 2014)) using bwa mem (Li 2013) with default settings. HTseq version 5.5 (Anders et al. 2015) was used to sum read counts for each gene. Differential expression analysis was performed using the program edgeR v.3.8.6. (Robinson et al. 2010) using a linear model to simultaneously analyze all twelve samples across two environments, with strain background and media type as factors and replicate samples paired. Data were normalized for visualization using the reads per kilobase per million mapped reads (RPKM) method. Hierarchical clustering analysis was performed using the R package Mclust (Scrucca et al. 2016) using model ‘VII’. Visualization was performed with the program Java Treeview (http://jtreeview.sourceforge.net/) (Saldanha 2004). Where indicated, data were clustered based on the RPKM values normalized to the mean value for each transcript across all strains (‘mean centered’). Functional enrichment analysis was performed by in-house scripts for the hypergeometric test using four different datasets previously defined (Chasman et al. 2014) or using FunSpec (Robinson et al. 2002, Boyle et al. 2004).
Data availability
All sequencing data are available in the NIH GEO database under accession number GSE118069. Strains used in this study and initial phenotyping are available in Table S1. Log2(fold change) expression data shown in figures are avialable in the following supplemental files: Figure 2A: Dataset 1; Figure 2B: Dataset 2; Figure 3: Dataset 3; Figure 4: Dataset4. Supplemental material available at Figshare: https://doi.org/10.25387/g3.7152551.
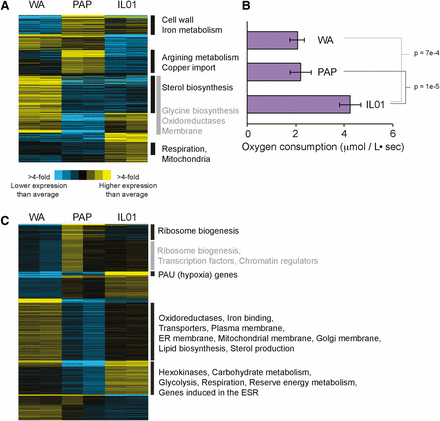
Strain-specific transcriptomic differences vary with oxygen. (A) Genes (rows) were clustered based on the mean-centered log2 RPKM. Shown are 474 genes whose expression was significant different (FDR < 1%) in WA, PAP, or IL01 compared to the mean expression of the three strains. (B) Rates of oxygen consumption during aerobic YPD growth. Asterisk indicates significant differences compared to the other strains. (C) Shown are 1,923 differentially expressed in each strain compared to the mean (FDR < 1%), as described in (A). Functional enrichments (P < 1e-5) are listed for each group; gray boxes are used simply for demarcation.
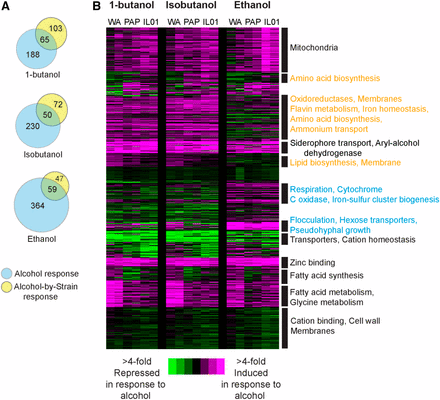
Expression responses to butanol, isobutanol, and ethanol in aerobic conditions. (A) Venn diagrams represent the number of genes whose expression responds to alcohol independent of strain and genes with a strain-specific response, for each alcohol. (B) Hierarchical clustering of 776 genes identified as differentially expressed in response to 0.8% butanol, 1% isobutanol, or 4% ethanol compared to YPD, in cells growing aerobically. Genes were clustered based on the log2(fold change) in expression in each strain (WA, PAP, IL) grown in the indicated alcohol vs. YPD, in biological duplicate. Expression changes are colored to indicate induciton (pink) and repression (green) according to the key. Functional categories enriched in each cluster (P < 1e-4, FunSpec) are shown to the right of each cluster. Responses specific to ethanol are highlighted in blue text and those specific to butanols are highlighted in orange text.
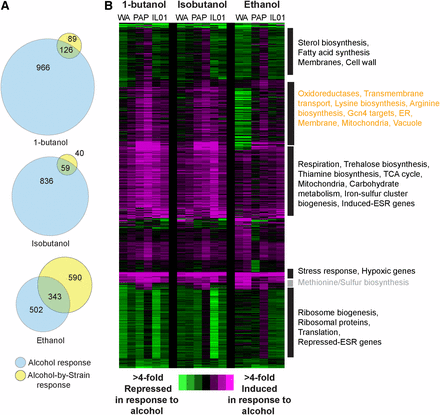
Expression responses to butanol, isobutanol, and ethanol under anaerobic conditions. (A) Venn diagrams represent the number of genes whose expression responds to alcohol independent of strain and genes with a strain-specific response, for each alcohol treatment administred anaerobicaly. (B) Hierarchical clustering of 2,075 genes differentially expressed in response to 0.8% butanol, 1% isobutanol, or 3% ethanol compared to YPD, under anaerobic conditions, as described in Figure 3. We note that the expression values for alcohol-treated cells were slightly higher for all strains in the second replicate, a feature that was accounted for in the replicate-paired statistical analysis.
Results
Alcohol tolerance varies Across Saccharomyces cerevisiae strains and with oxygen availability
To characterize variation in alcohol tolerance, we first surveyed 165 strains of Saccharomyces cerevisiae collected from a variety of geographical locations and ecological niches for their ability of cells to grow in 1-butanol (Table S1, see Methods). 1-butanol tolerance is clearly a quantitative trait, and we observed a wide range of tolerances spanning fourfold variation in final cell density of the most sensitive and tolerant strains (Figure 1A). Based on these phenotypes, we chose three strains with different degrees of tolerance (and that also produce viable spores, to support future genetic mapping studies) for more detailed investigation: a West African strain isolated from bili wine, NCYC3290 (WA), a mosaic strain representing admixed pure lineages that was isolated from a rotten papaya Y7568 (PAP) and a mosaic strain isolated from soil IL01 (IL) (Table S1).
We measured growth of our selected strains in rich laboratory medium (YPD) and in medium supplemented with 1% 1-butanol, 1.2% isobutanol, and 5% ethanol, under aerobic (Figure 1B) and anaerobic conditions (Figure 1C). Interestingly, anaerobiosis affected strains in different ways. IL01 followed by PAP were most tolerant to butanols, whereas both IL01 and PAP showed similar aerobic tolerance to ethanol. WA was the most sensitive to all alcohols but showed particularly poor growth in ethanol (Figure 1A). However, the strains showed different phenotypes under anaerobic growth: in the absence of alcohols, PAP was the fastest growing strain and grew to nearly the same final cell density as in aerobic conditions, unlike the other strains that grew slower anaerobically. All of the strains were more sensitive to alcohols anaerobically, and the differences in tolerance to this dose of alcohol were minimal under anaerobic conditions (although WA was unable to survive higher doses of ethanol anaerobically). Thus, the results show an interesting genotype (strain) by environment (oxygen availability) by environment (alcohol exposure) interaction, such that strains show significant differences in ranked tolerance depending on the alcohol and the oxygen status.
Strain-specific transcriptomic responses to anaerobic conditions
The condition-specific differences in strain performance provided an opportunity to investigate differences in tolerance mechanisms, with and without available oxygen. We first analyzed transcriptomes of the three strains grown in rich YPD medium with and without oxygen. There were 474 differentially expressed genes (FDR <1%) across the three strains grown aerobically (Figure 2A), and hierarchical clustering revealed groups of functionally related genes (Dataset 1). Interestingly, one cluster that was expressed higher in the fastest growing IL01 was significantly enriched for genes involved in aerobic respiration (Figure 2A), raising the possibility that IL01 relies more on respiration than the other studied strains even in the presence of glucose. To test this, we measured oxygen consumption during aerobic growth on rich medium. Indeed, IL01 consumed oxygen at a significantly faster rate compared to WA and PAP (Figure 2B), which could contribute both to its faster growth rate and improved alcohol tolerance specifically in aerobic conditions (see Discussion).
Next, we investigated transcriptome differences under anaerobic conditions in the absence of added alcohols: the number of differentially expressed genes across strains was over three times greater than under aerobic conditions at 1,923 genes (FDR < 1%, Figure 2C, Dataset 2). Interestingly, it was the slowest growing strain WA that showed high expression of many genes, enriched for those related to membrane, ergosterol, and fatty-acid synthesis (Figure 2C). This was intriguing, because WA was the slowest growing strain under anaerobic conditions (predicted to impact membrane fluidity) and the most sensitive to membrane-targeting alcohols (see Discussion). IL01 showed significantly higher expression of genes related to energy production, mobilization of energy reserves, gene induced in the environmental stress response, and PAU genes that are induced as part of the hypoxic response (Rachidi et al. 2000, Luo & van Vuuren 2009), whereas the fastest anaerobic grower PAP displayed the lowest expression of these genes (see Discussion).
Transcriptome responses to aerobic alcohol exposure implicate common and alcohol-specific responses
To investigate mechanisms of alcohol tolerance, we next compared the transcriptome responses to 0.8% 1-butanol, 1% isobutanol, and 4% ethanol in strains growing under aerobic conditions. These doses are higher than titers produced by engineered strains, but were chosen to provoke a robust stress response but allow all three strains to grow. We used a linear model to identify genes with whose expression was responsive to alcohol independent of strain background and genes affected by a strain-by-media (i.e., gene-by-environment) interaction (see Methods). We identified hundreds of genes whose expression responded to each alcohol, and many genes with strain-by-alcohol interactions (Figure 3). We hierarchically clustered the combined set of 776 genes that responded to any of the three alcohols (including those with strain-specific alcohol responses, Figure 3A, Dataset 3).
Many of the expression responses were similar across strains and regardless of alcohol identity. This included induction of genes involved in mitochondrial functions, siderophore transport, aryl-alcohol dehydrogenases, and zinc binding, and repression of genes encoding transporters and many other membrane proteins (Figure 3B). In contrast, there were also several responses unique to 1-butanol and isobutanol compared to ethanol. Butanol isoforms induced a different set of genes linked to membrane synthesis, as well as genes associated with oxidoreduction, iron homeostasis, amino-acid biosynthesis, and ammonium transport. Interestingly, ethanol uniquely triggered in all three strains the induction of genes involved in flocculation, hexose transport, pseudohyphal growth, and respiration, whereas butanol isoforms did not (Figure 3B). These responses are also seen in cells undergoing filamentous growth, and ethanol is known to trigger the response in some strains (Lorenz et al. 2000). There were also several aspects of the responses that were different across strains. Most notably, WA showed much stronger induction of lipid and fatty-acid biosynthesis genes, especially in response to butanols, again suggesting that this strain may experience specific defects in membrane function (see Discussion).
Strain-specific responses to alcohol are different anaerobically
Most industrial fermentations are anaerobic, however the majority of studies analyzing alcohol toxicity are based on aerobic conditions. We therefore analyzed the transcriptome response to each alcohol anaerobically, comparing the response to 0.8% 1-butanol, 1% isobutanol, and 3% ethanol to anaerobic YPD growth. Pooled together, the analysis identified 2,075 genes that were differentially expressed in response to alcohols in one or more strains (Dataset 4). All three alcohols were more inhibitory in anaerobic conditions (see Figure 1), and we correspondingly observed 2.sixfold more genes differentially regulated in response to alcohols under anaerobic vs. aerobic conditions (Figure 4). Many of the responses were common to all three alcohols; however, we observed one gene cluster with a specific response to butanols (Figure 4B, orange text). This group of induced genes was enriched for genes related to membranes, mitochondrial function, and Gcn4 targets involved in amino acid biosynthesis. These genes were either not strongly induced or in the case of WA were strongly repressed in response to ethanol. WA had other unique responses to ethanol and likely contributed to the large number of strain-by-ethanol responses (Figure 4A). In contrast, the anaerobic response to butanols was largely similar across the three strains.
To better understand the differences in alcohol responses, independent of strain-specific responses, we compared the alcohol-responsive genes identified by each linear model (FDR < 1%) when cells were grown with and without oxygen (Figure 5) and investigated functional enrichment among genes in each set. This provides a complementary analysis to the clustering described above. Anaerobically, all three alcohols triggered the induction of genes related to energy metabolism. These include genes involved in respiration, TCA cycle, energy reserve metabolism, and/or the stress response, and together suggests energy limitation during anaerobic alcohol defense (see Discussion). Aerobically, butanols induced genes involved in iron homeostasis, oxidoreduction, and amino-acid biosynthesis, with isobutanol producing a broader effect across Gcn4 targets as well as other genes related to lysine, arginine, and purine biosynthesis (see also Figure 4). Butanols triggered reduced expression of respiration genes during aerobic growth but induced expression anaerobically – this was in contrast to ethanol that led to the induction of respiration genes both aerobically and anaerobically (Figure 3-5). In at least two of the three strains, ethanol also triggered reduced expression of genes encoding plasma membrane and cell surface markers (Figure 5, 4B) that were not strongly triggered by butanols despite their higher toxicity. These results suggest cellular targets and/or secondary effects of the different alcohols and mechanisms cells use to respond.
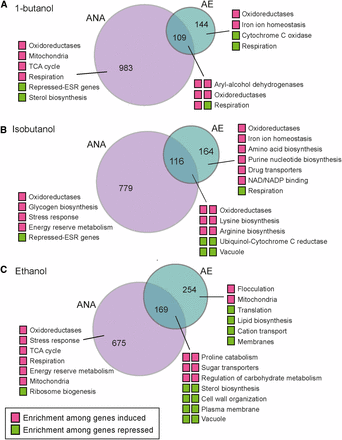
Comparison of aerobic and anaerobic alcohol responses. Each Venn diagram compares genes differentially expressed (FDR < 1%) under aerobic (AE), anaerobic (ANA), or both conditions for (A) 1-butanol, (B) isobutanol, and (C) ethanol. Colored boxes represent genes induced (magenta) or repressed (green) under those conditions; for genes in the overlap, the left box indicates anaerobic conditions and the right box represents aerobic conditions. Functional categories enriched for each gene group (P < 1e-4, FunSpec) are shown.
Discussion
With the pursuit of more energy-intensive renewable fuels, recent efforts have been focused on microbial production of butanol isoforms using both synthetic and metabolic engineering strategies (Dürre, 2011, Buijs et al. 2013). While butanols are expected to target similar cellular processes as the better studied ethanol, the unique features of butanol stress responses are not well understood. Our comparative transcriptomic response has generated new insights into how genetics and environment influence the cellular response to these alcohols.
While we set out to study alcohol responses by exploring natural diversity in tolerances, an important result of our study is that different strains of S. cerevisiae respond differently to anaerobic growth in the absence of added stress. We were surprised to find growth properties and relative alcohol sensitivities would differ across strains depending on oxygen availability. IL01 grew on par with the PAP strain aerobically but grew significantly slower anaerobically (Figure 1); that this strain respires at a higher rate than other strains aerobically (Figure 2B) suggests that it relies more on respiration for energy generation, and hence suffers a disadvantage when respiration is blocked by anaerobiosis. In the absence of added alcohols, the WA strain showed uniquely high expression of genes enriched for membrane functions, sterol synthesis, and fatty acid production, both aerobically and anaerobically. Many of these genes were repressed in WA responding to alcohols, especially ethanol anaerobically (Figure 4). Since membrane integrity and fluidity are primary targets of alcohols, we suggest that the increased sensitivity of the WA strain may be due to an underlying difference in membrane composition or the ability to maintain it. These results highlight the utility of incorporating a comparative genomic approach to studying the response to industrial stresses that can vary significantly across strains. It will be important to characterize strains that have been engineered for high butanol production, including naturally tolerant strains being engineered for isobutanol tolerance.
Our results also point to commonalities and unique features in the responses to different alcohols across multiple strains. All strains were more sensitive to alcohols anaerobically, and consequently all strains showed more alcohol-responsive expression changes under these conditions. The common induction of genes related to energy metabolism is consistent with the idea of energy shortages during anaerobic alcohol defense. Genes involved in respiration and alternate energy mobilization are commonly induced during stress even in the absence of oxygen (Gasch et al. 2000, Lahtvee et al. 2016, Malina et al. 2018). Furthermore, up-regulation of respiration proteins was associated with evolved 1-butanol tolerance in yeast (Ghiaci et al. 2013). Yet, here we found that both 1-butanol and isobutanol treatment led to reduced aerobic expression of respiration genes. The reason for this is unclear, but could be related to other mitochondrial processes influenced by butanol response, including NADH/NAD+ rebalancing or iron-sulfur cluster generation.
Prior investigation of butanol responses in yeast and bacteria pointed to signatures of protein misfolding and oxidative stress, involving protein-folding chaperones, proteasome-dependent protein degradation, and redox rebalancing (Rutherford et al. 2010, Ghiaci et al. 2013, González-Ramos et al. 2013, Crook et al. 2016). We observed multiple protein folding chaperones induced by butanols under anaerobic stress, but we did not see induction of proteasome genes as a group (Datasets 3, 4). Many oxidoreductases, including alcohol and aldehyde dehydrogenases (including ADH5, ALD5, GDH1, MDH2, DLD1) were induced uniquely by butanols, while others that are part of the yeast Environmental Stress Response (Gasch et al. 2000) were induced by all alcohols. Several of these oxidoreductases may represent specific detoxification mechanisms against butanols. Finally, 1-butanol and especially isobutanol led to the induction of many genes involved in amino acid biosynthesis, including biosynthetic genes regulated by the transcription factor Gcn4. Gcn4 can be activated by amino acid starvation but also uncharged tRNAs or translation defects (Hinnebusch 2005). It is unclear why the response is induced here, but alcohols are known to inhibit translation elongation (Ashe et al. 2001, Haft et al. 2014). It is also possible that the response is triggered by isobutanol itself, produced as a byproduct of valine biosynthesis; however, we saw no notable expression changes of ILV genes involved in that pathway.
Our results also raise important considerations about genotype-phenotype relationships and how they change in different environments. Both growth properties and gene expression varied dependent on strains as well as the presence of oxygen, alcohols, or both conditions, revealing important strain-by-oxygen-by-alcohol interactions. While genotype-by-environment effects are well appreciated in quantitative genetics, combinatorial effects of more complex environmental changes are generally less well studied. Our results set the stage for further investigation of genotype-by-environment-by-environment changes and how they can be leveraged for understanding and engineering industrially relevant traits.
Acknowledgments
We thank M. Place for bioinformatic assistance. MS was supported by a predoctoral fellowship from the NSF. Work was funded by the Great Lakes Bioenergy Research Center, U.S. Department of Energy, Office of Science, Office of Biological and Environmental Research under Award Numbers DE-SC0018409 and DE-FC02-07ER64494.
Footnotes
Supplemental material available at Figshare: https://doi.org/10.25387/g3.7152551.
Communicating editor: B. Andrews
Literature Cited
Author notes
Present address: Cargill, Inc.
Present address: Gene-by-Gene, Houston, TX 77008

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}